地学前缘 ›› 2020, Vol. 27 ›› Issue (6): 255-275.DOI: 10.13745/j.esf.sf.2020.6.13
苏新1( ), 曲莹1,2, 陈芳3,4, 杨胜雄3,4, 周洋3,4, 崔鸿鹏1, 于翀涵1, 滕田田1
), 曲莹1,2, 陈芳3,4, 杨胜雄3,4, 周洋3,4, 崔鸿鹏1, 于翀涵1, 滕田田1
收稿日期:2020-03-19
修回日期:2020-05-18
出版日期:2020-11-02
发布日期:2020-11-02
作者简介:苏 新(1957—),女,教授,地层古生物和海洋地质专业。E-mail: xsu@cugb.edu.cn
基金资助:
SU Xin1(), QU Ying1,2, CHEN Fang3,4, YANG Shengxiong3,4, ZHOU Yang3,4, CUI Hongpeng1, YU Chonghan1, TENG Tiantian1
Received:2020-03-19
Revised:2020-05-18
Online:2020-11-02
Published:2020-11-02
摘要:
为探讨冷泉区底栖有孔虫组合特征、受控因素及其冷泉微生境随时间的变迁,本文对台西南海盆取自九龙甲烷礁和海洋四号区冷泉区的973-4和973-5两根岩心展开了底栖有孔虫及其壳体氧碳同位素研究。测年结果揭示两根岩心为晚更新世约5万年来海洋氧同位素期(MIS)MIS 3至MIS 1早期的沉积序列。两个站位共识别了底栖有孔虫79属233种,优势类别在973-4组合中为Uvigerina(23.3%)、Bulimina(10.71%)和Cibicidoides(9.87%),在973-5组合中是Bulimina(20.6%),两站位的组合优势和常见属种均以内生类别为主。有孔虫分异度显然同时受到正常深海环境因子TOC和沉积物粒度的影响。总体上,优势和常见类群与TOC相关性较弱,但与δ18OUvigerina spp.有不同程度的相关性,说明有孔虫还受冷泉特殊营养物质和流体因子影响。5万年来,973-4和973-5站位底栖有孔虫组合生活的冷泉微生境,经历了由双壳-自生碳酸盐岩(MIS 3至MIS 2早期)向双壳-菌席(MIS 2晚期至MIS 1早期)的变迁。底栖有孔虫的优势类群也随微生境的变迁而演替,如973-4站位MIS 3-MIS 1的优势类群依次为U.peregrina、Cibicidoides-Bulimina、U.vadescens和Cibicides,973-5站位为Chilostomella+ Globobulimina、Cibicidoides、Bulimina。有孔虫壳体氧碳同位素特征也随时间改变,从MIS 3到MIS 2早期,在自生碳酸盐岩水岩交换背景下,具有富δ18O和亏损δ13C特点(3.5‰~4.49‰,-2‰~-0.2‰); MIS 2晚期—MIS 1早期因双壳和菌席的生物地球化学作用影响,具有略富集δ18O和略微亏损δ13C的特征(2.5‰~3.5‰,-1‰~-0.1‰)。自5万年来两个区甲烷渗漏逐渐减弱,其间发生了几次增强事件。973-4站位记录了1次持续时间约10 ka的增强事件(35~25 ka);973-5站位记录3次(45 ka,35 ka,14~12 ka)。其中,45 ka时海底上涌的甲烷通量可能最大,在海底表面形成水合物。35 ka时的甲烷喷溢增强事件可能为区域性事件。
中图分类号:
苏新, 曲莹, 陈芳, 杨胜雄, 周洋, 崔鸿鹏, 于翀涵, 滕田田. 台西南深海底栖有孔虫及其5万年来冷泉微生境变迁记录[J]. 地学前缘, 2020, 27(6): 255-275.
SU Xin, QU Ying, CHEN Fang, YANG Shengxiong, ZHOU Yang, CUI Hongpeng, YU Chonghan, TENG Tiantian. Deep sea benthic foraminifera from the Taixinan Basin and changes of their cold seep microhabitats during the past 50000 years[J]. Earth Science Frontiers, 2020, 27(6): 255-275.
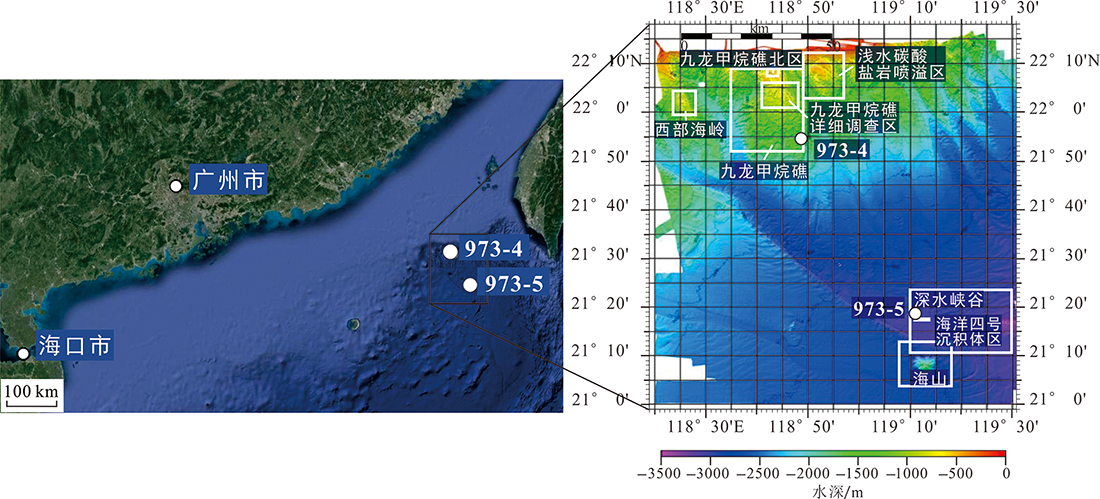
图1 973-4和973-5站位地理位置以及九龙甲烷礁和海洋四号冷泉区范围(右侧海底图据文献[13]修改)
Fig.1 Geographical location of Cores 973-4 and 973-5 and distributions of the Jiulong Methane Reef and Haiyang No.4 cold seep areas (bathymetric chart modified after [13])

图2 九龙甲烷礁部分典型冷泉地貌表征和微生境 A:厚层自生碳酸盐岩、裂隙及裂隙壁的菌席[10,13-14];B:自生碳酸盐岩和活双壳群落生境区[10]。
Fig.2 Parts of typical cold seep manifestations and microhabitats on the Jiulong Methane Reef

图3 海洋四号沉积体区部分典型冷泉地貌表征和微生境 A:较丰富的双壳群落(死亡)和菌席(灰白色斑片状)混杂区[14];B:小斑块状分布的双壳群落(死亡)和周边沉积物。
Fig.3 Selected typical cold seep manifestations and microhabitats in the Haiyang No.4 cold seep area
| 保压取心 重力柱 | 水深 /m | 岩心 总长/m | 冷泉区 | 已知冷泉区典型 海底地貌特征 | 站位其他地理信息 | 冷泉沉积物岩心 主要标识特征 | 有孔虫 样品信息 |
|---|---|---|---|---|---|---|---|
| 973-4 | 1 660 | 13.65 | 九龙 甲烷礁 | 广布自生碳酸盐礁体、礁体裂隙间发育白色菌席 | 九龙甲烷礁体海脊南部斜坡 | H2S气味,H2S浸染黑斑,低部甲烷高含量 | 取自27~770 m岩心段,间距20 cm,31个样,干样10 g |
| 973-5 | 2 998 | 9.35 | 海洋四号 沉积体 | 斑状分布的密集死亡双壳群落 | 台湾海底峡谷中段 | H2S气味,H2S浸染黑斑,低部甲烷高含量 | 取自28~935 m岩心段,间距15 cm,53个样,干样5 g |
表1 973-4和973-5岩心概况及冷泉环境标识特征
Table 1 General information of Cores 973-4 and 973-5 and their cold seep environmental identification characteristics
| 保压取心 重力柱 | 水深 /m | 岩心 总长/m | 冷泉区 | 已知冷泉区典型 海底地貌特征 | 站位其他地理信息 | 冷泉沉积物岩心 主要标识特征 | 有孔虫 样品信息 |
|---|---|---|---|---|---|---|---|
| 973-4 | 1 660 | 13.65 | 九龙 甲烷礁 | 广布自生碳酸盐礁体、礁体裂隙间发育白色菌席 | 九龙甲烷礁体海脊南部斜坡 | H2S气味,H2S浸染黑斑,低部甲烷高含量 | 取自27~770 m岩心段,间距20 cm,31个样,干样10 g |
| 973-5 | 2 998 | 9.35 | 海洋四号 沉积体 | 斑状分布的密集死亡双壳群落 | 台湾海底峡谷中段 | H2S气味,H2S浸染黑斑,低部甲烷高含量 | 取自28~935 m岩心段,间距15 cm,53个样,干样5 g |

图4 973-5岩心异常沉积现象 A:456.5 cm处不整合界面;B:不整合面夹层中沉积物冲洗后照片,包括分选和磨圆程度高的滨浅海石英等碎屑,含少量双壳类碎屑;C:785 cm处的稀汤沉积(疑似水合物分解);D,E:自生碳酸盐岩;D:710 cm处自生碳酸盐岩碎屑; E:875~ 915 cm未固结或半固结泥质自生碳酸盐岩(红色箭头处)和贝壳碎片(黄色箭头处)。
Fig.4 Records of abnormal sedimentary in Core 973-5
| 岩心 | 平均 深度/cm | 测试结果14C 年龄/a B.P. | 深度控制点 年龄/ka | 岩心 | 平均 深度/cm | 测试结果14C 年龄/a B.P. | 深度控制点 年龄/ka |
|---|---|---|---|---|---|---|---|
| 973-4 | 0.0 | 9.64* | 973-5 | 0.0 | 7.84 | ||
| 973-4 | 15.0 | 9.95* | 973-5 | 17.5 | 8 428±34 | 8.43 | |
| 973-4 | 102.5 | 11 766±72 | 11.77 | 973-5 | 105.5 | 11 337±70 | 11.34 |
| 973-4 | 202.5 | 13 838±69 | 13.84 | 973-5 | 207.5 | 13 315±50 | 13.32 |
| 973-4 | 302.5 | 17 405±228 | 17.41 | 973-5 | 254.0 | 15 721±187 | 15.72 |
| 973-4 | 448.5 | 18 990±59 | 18.99 | 973-5 | 376.5 | 18 866±56 | 18.87 |
| 973-4 | 477.5 | 21 771±202 | 21.77 | 973-5 | 456.5 | 19 867±70 | 19.87 |
| 973-4 | 484.5 | 22 599±127 | 22.60 | 973-5 | 460.0 | 34 630±200 | 31.74* |
| 973-4 | 602.5 | 36 245±230 | 36.25 | 973-5 | 526.5 | 34 146±210 | 34.15 |
| 973-4 | 770.0** | 55.62* | 973-5 | 626.5 | 37 772±280 | 37.77 | |
| 973-5 | 935.0 | 48.77* |
表2 973-4和973-5岩心所获得的AMS14C测年结果及深度控制点年龄
Table 2 AMS14C dating results and chronological framework for Cores 973-4 and 973-5
| 岩心 | 平均 深度/cm | 测试结果14C 年龄/a B.P. | 深度控制点 年龄/ka | 岩心 | 平均 深度/cm | 测试结果14C 年龄/a B.P. | 深度控制点 年龄/ka |
|---|---|---|---|---|---|---|---|
| 973-4 | 0.0 | 9.64* | 973-5 | 0.0 | 7.84 | ||
| 973-4 | 15.0 | 9.95* | 973-5 | 17.5 | 8 428±34 | 8.43 | |
| 973-4 | 102.5 | 11 766±72 | 11.77 | 973-5 | 105.5 | 11 337±70 | 11.34 |
| 973-4 | 202.5 | 13 838±69 | 13.84 | 973-5 | 207.5 | 13 315±50 | 13.32 |
| 973-4 | 302.5 | 17 405±228 | 17.41 | 973-5 | 254.0 | 15 721±187 | 15.72 |
| 973-4 | 448.5 | 18 990±59 | 18.99 | 973-5 | 376.5 | 18 866±56 | 18.87 |
| 973-4 | 477.5 | 21 771±202 | 21.77 | 973-5 | 456.5 | 19 867±70 | 19.87 |
| 973-4 | 484.5 | 22 599±127 | 22.60 | 973-5 | 460.0 | 34 630±200 | 31.74* |
| 973-4 | 602.5 | 36 245±230 | 36.25 | 973-5 | 526.5 | 34 146±210 | 34.15 |
| 973-4 | 770.0** | 55.62* | 973-5 | 626.5 | 37 772±280 | 37.77 | |
| 973-5 | 935.0 | 48.77* |
| 氧同位素分期界限 | 年龄*/ka | 973-4岩心深度/cm | 973-5岩心深度/cm |
|---|---|---|---|
| MIS 1/MIS 2 | 14 | 207.04 | 220.74 |
| MIS 2/MIS 3 | 29 | 539.85 | 459.19 |
| MIS 3/MIS 4 | 57 |
表3 973-4和973-5岩心氧同位素分期界限的深度和年龄
Table 3 Depths and ages of oxygen isotope stage boundaries for Cores 973-4 and 973-5
| 氧同位素分期界限 | 年龄*/ka | 973-4岩心深度/cm | 973-5岩心深度/cm |
|---|---|---|---|
| MIS 1/MIS 2 | 14 | 207.04 | 220.74 |
| MIS 2/MIS 3 | 29 | 539.85 | 459.19 |
| MIS 3/MIS 4 | 57 |
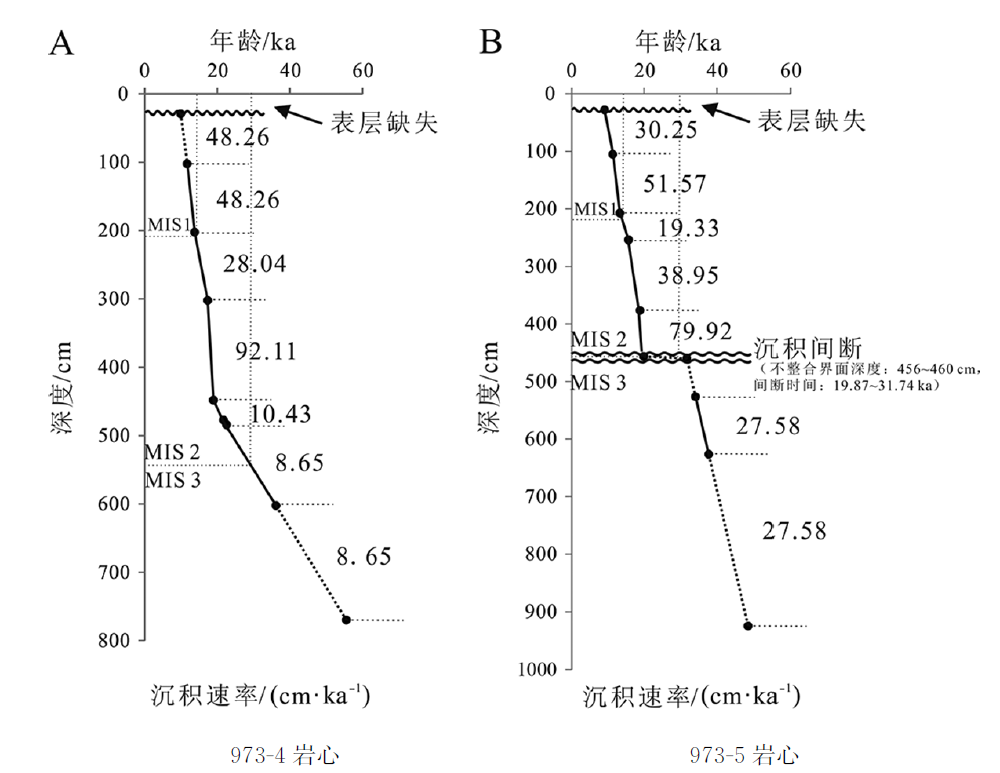
图5 973-4岩心(A)和973-5岩心(B)的年龄与深度对比及沉积速率变化
Fig.5 Age and depth correlation and variation in sedimentary rate for Cores 973-4 (A) and 973-5 (B)
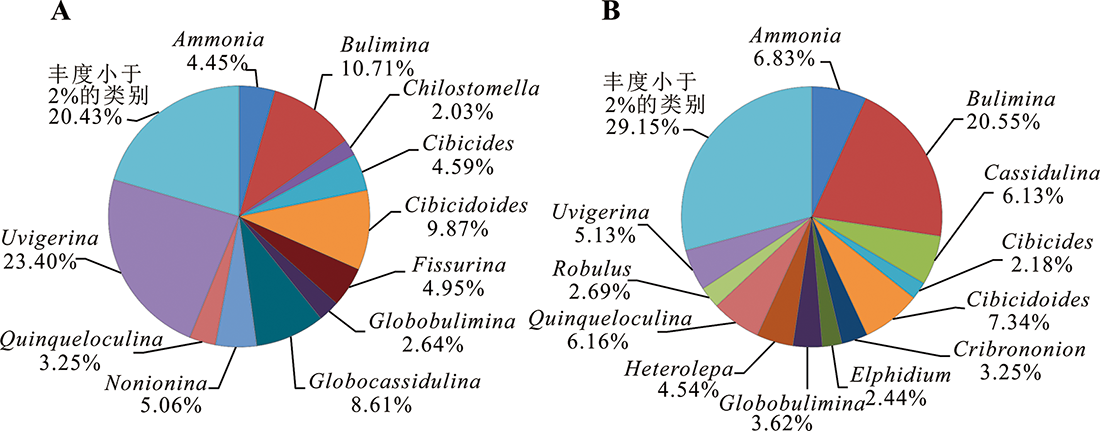
图6 973-4(A)和973-5(B)岩心底栖有孔虫按平均含量大于2%的属归纳的组合特征
Fig.6 Characteristics of benthic foraminiferal assemblages in Cores 973-4 and 973-5, presenting separately only genera with an average abundance more than 2%
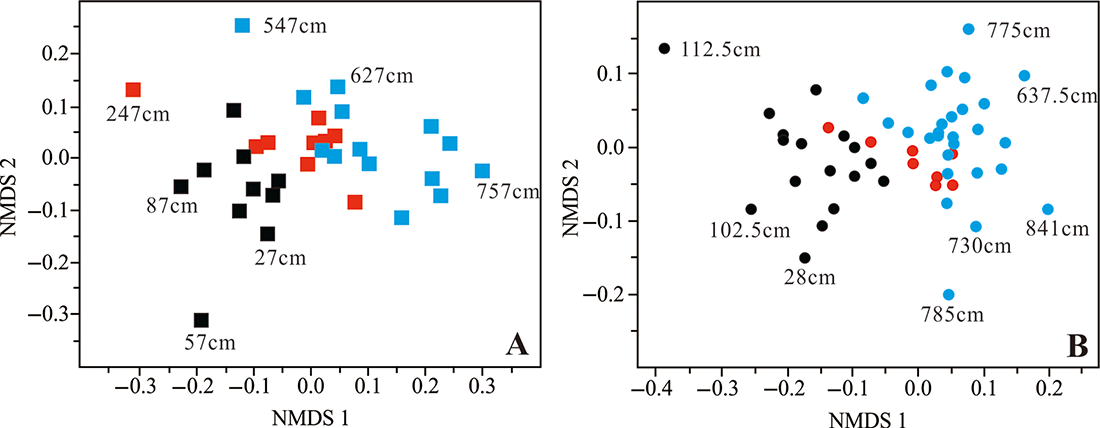
图7 基于Bray-Curtis距离的MIS 3-MIS 1时期主要有孔虫类群非度量多尺度(NMDS)分析 A:973-4岩心,B:973-5岩心;图中点为各深度样有孔虫综合信息(属种丰度);黑、红、蓝色分别代表MIS 1、MIS 2和MIS 3时期的样品;数字为部分样品深度。
Fig.7 Non metric multiscale (NMDS) analysis of major foraminifera taxa during MIS 3-MIS 1 based on Bray-Curtis distance
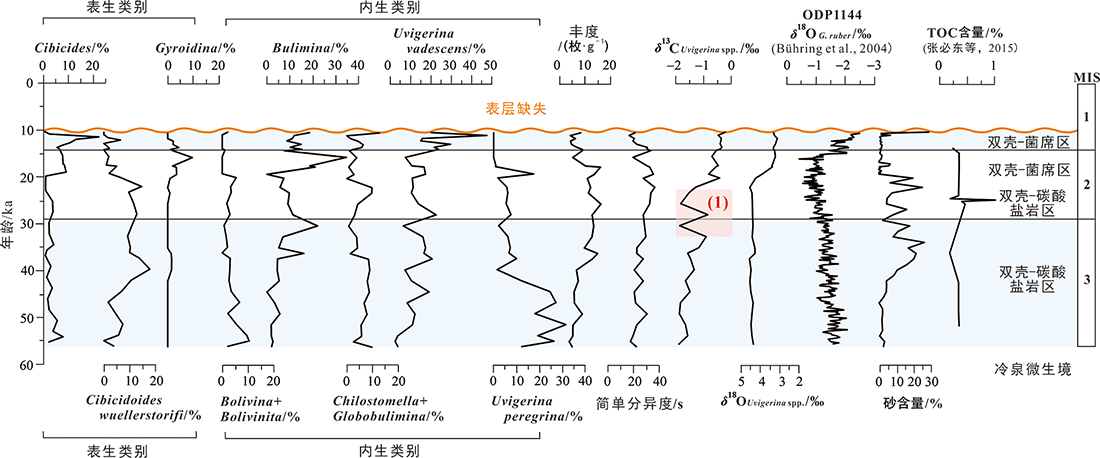
图8 MIS 3-MIS 1期973-4岩心底栖有孔虫主要类别含量、有孔虫丰度和分异度及其与氧碳同位素、砂和TOC含量对比(ODP 1144站位δ18OG.ruber记录引自文献[46],TOC含量数据引用文献[47]) 图中(1)和箭头标识为甲烷渗流活动增强时段(见本文5.5讨论部分)。
Fig.8 Comparisons of abundance of major benthic foraminifera taxa, fauna abundance and diversity with oxygen/carbon isotope, sand and TOC contents for Core 973-4 during MIS 3-MIS 1 period (record of δ18 O G . ruber at Site ODP1144 adapted from [46]; TOC data adapted from [47])
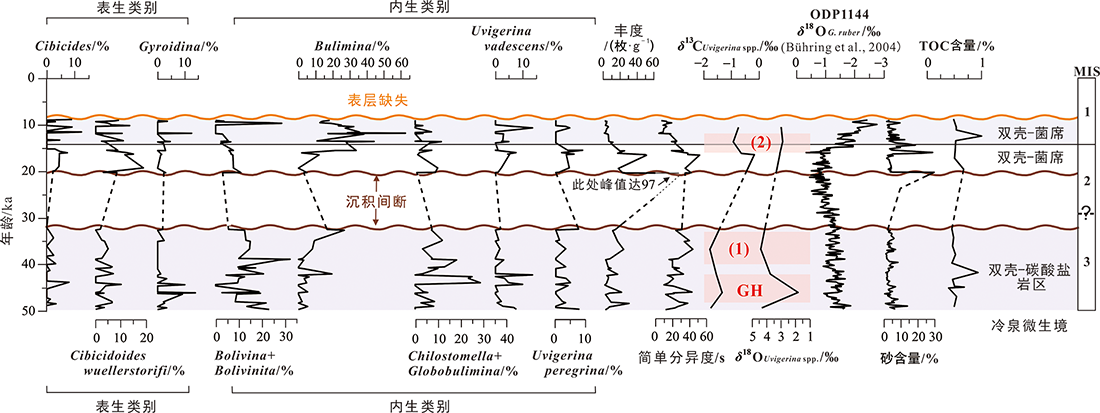
图9 MIS 3-MIS 1时期973-5岩心底栖有孔虫主要类别含量、有孔虫丰度和分异度及其与氧碳同位素、砂和TOC含量对比(ODP1144站位δ18OG.ruber记录引自文献[46]) 图中(1)、(2)和GH标识为指示甲烷渗流活动增强和水合物形成事件(见本文5.5讨论部分)。
Fig.9 Comparisons of abundance of major benthic foraminifera taxa, fauna abundance and diversity with oxygen/carbon isotope, sand and TOC contents for Core 973-5 during MIS 3-MIS 1 period (record of δ18 O G . ruber at ODP Site 1144 adapted from [46])
| 分样号 | 深度/cm | 年龄/ka | 有孔虫属 | δ13C/‰ | δ18O/‰ | 分样号 | 深度/cm | 年龄/ka | 有孔虫属 | δ13C/‰ | δ18O/‰ |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 973-4-1 | 25 | 10.16 | Uvigerina | -0.21 | 3.25 | 973-4-57 | 597 | 35.61 | Uvigerina | -1.56 | 4.20 |
| 973-4-4 | 57 | 10.82 | Uvigerina | -0.47 | 3.20 | 973-4-58 | 607 | 36.77 | Uvigerina | -1.46 | 4.49 |
| 973-4-7 | 87 | 11.45 | Uvigerina | -0.36 | 3.05 | 973-4-60 | 627 | 39.08 | Uvigerina | -1.33 | 4.41 |
| 973-4-13 | 147 | 12.69 | Uvigerina | -0.40 | 3.15 | 973-4-62 | 647 | 41.39 | Uvigerina | -1.64 | 4.47 |
| 973-4-15 | 177 | 13.31 | Uvigerina | -0.34 | 3.25 | 973-4-64 | 667 | 43.70 | Uvigerina | -1.48 | 4.42 |
| 973-4-17 | 197 | 13.72 | Uvigerina | -0.39 | 3.25 | 973-4-66 | 687 | 46.02 | Uvigerina | -1.82 | 4.3 |
| 973-4-18 | 207 | 14.00 | Uvigerina | -0.54 | 3.19 | 973-4-68 | 707 | 48.33 | Uvigerina | -1.86 | 4.36 |
| 973-4-21 | 243 | 15.28 | Uvigerina | -0.69 | 3.15 | 973-4-70 | 727 | 50.64 | Uvigerina | -1.64 | 4.41 |
| 973-4-27 | 297 | 17.21 | Uvigerina | -0.47 | 3.33 | 973-4-72 | 747 | 52.96 | Uvigerina | -1.56 | 4.36 |
| 973-4-28 | 307 | 17.45 | Uvigerina | -0.55 | 3.31 | 973-4-73 | 763 | 54.81 | Uvigerina | -1.89 | 4.28 |
| 973-4-42 | 447 | 18.97 | Uvigerina | -0.81 | 3.84 | 973-5-8 | 70 | 10.16 | Uvigerina | -0.72 | 2.97 |
| 973-4-43 | 457 | 19.81 | Uvigerina | -0.42 | 4.17 | 973-5-32 | 205 | 13.27 | Uvigerina | -0.91 | 2.86 |
| 973-4-45 | 477 | 21.72 | Uvigerina | -1.27 | 4.47 | 973-5-42 | 253 | 15.68 | Uvigerina | -0.62 | 3.05 |
| 973-4-46 | 487 | 22.89 | Uvigerina | -1.51 | 4.36 | 973-5-44 | 263 | 15.95 | Uvigerina | -0.15 | 3.06 |
| 973-4-48 | 507 | 25.20 | Uvigerina | -1.81 | 4.34 | 973-5-98 | 577 | 35.98 | Uvigerina | -1.75 | 4.38 |
| 973-4-50 | 527 | 27.51 | Uvigerina | -0.84 | 4.33 | 973-5-124 | 723 | 41.27 | Uvigerina | -1.52 | 3.72 |
| 973-4-52 | 547 | 29.83 | Uvigerina | -1.85 | 4.31 | 973-5-137 | 830 | 45.15 | Uvigerina | -1.31 | 1.79 |
| 973-4-54 | 567 | 32.14 | Uvigerina | -0.90 | 4.35 | 973-5-150 | 920 | 48.42 | Uvigerina | -1.72 | 4.30 |
| 973-4-56 | 587 | 34.45 | Uvigerina | -1.16 | 4.21 |
表4 973-4和973-5岩心底栖有孔虫Uvigerina spp.壳体稳定氧碳同位素记录
Table 4 Stable oxygen and carbon isotope records of benthic foraminifera Uvigerina spp. shells in Cores 973-4 and 973-5
| 分样号 | 深度/cm | 年龄/ka | 有孔虫属 | δ13C/‰ | δ18O/‰ | 分样号 | 深度/cm | 年龄/ka | 有孔虫属 | δ13C/‰ | δ18O/‰ |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 973-4-1 | 25 | 10.16 | Uvigerina | -0.21 | 3.25 | 973-4-57 | 597 | 35.61 | Uvigerina | -1.56 | 4.20 |
| 973-4-4 | 57 | 10.82 | Uvigerina | -0.47 | 3.20 | 973-4-58 | 607 | 36.77 | Uvigerina | -1.46 | 4.49 |
| 973-4-7 | 87 | 11.45 | Uvigerina | -0.36 | 3.05 | 973-4-60 | 627 | 39.08 | Uvigerina | -1.33 | 4.41 |
| 973-4-13 | 147 | 12.69 | Uvigerina | -0.40 | 3.15 | 973-4-62 | 647 | 41.39 | Uvigerina | -1.64 | 4.47 |
| 973-4-15 | 177 | 13.31 | Uvigerina | -0.34 | 3.25 | 973-4-64 | 667 | 43.70 | Uvigerina | -1.48 | 4.42 |
| 973-4-17 | 197 | 13.72 | Uvigerina | -0.39 | 3.25 | 973-4-66 | 687 | 46.02 | Uvigerina | -1.82 | 4.3 |
| 973-4-18 | 207 | 14.00 | Uvigerina | -0.54 | 3.19 | 973-4-68 | 707 | 48.33 | Uvigerina | -1.86 | 4.36 |
| 973-4-21 | 243 | 15.28 | Uvigerina | -0.69 | 3.15 | 973-4-70 | 727 | 50.64 | Uvigerina | -1.64 | 4.41 |
| 973-4-27 | 297 | 17.21 | Uvigerina | -0.47 | 3.33 | 973-4-72 | 747 | 52.96 | Uvigerina | -1.56 | 4.36 |
| 973-4-28 | 307 | 17.45 | Uvigerina | -0.55 | 3.31 | 973-4-73 | 763 | 54.81 | Uvigerina | -1.89 | 4.28 |
| 973-4-42 | 447 | 18.97 | Uvigerina | -0.81 | 3.84 | 973-5-8 | 70 | 10.16 | Uvigerina | -0.72 | 2.97 |
| 973-4-43 | 457 | 19.81 | Uvigerina | -0.42 | 4.17 | 973-5-32 | 205 | 13.27 | Uvigerina | -0.91 | 2.86 |
| 973-4-45 | 477 | 21.72 | Uvigerina | -1.27 | 4.47 | 973-5-42 | 253 | 15.68 | Uvigerina | -0.62 | 3.05 |
| 973-4-46 | 487 | 22.89 | Uvigerina | -1.51 | 4.36 | 973-5-44 | 263 | 15.95 | Uvigerina | -0.15 | 3.06 |
| 973-4-48 | 507 | 25.20 | Uvigerina | -1.81 | 4.34 | 973-5-98 | 577 | 35.98 | Uvigerina | -1.75 | 4.38 |
| 973-4-50 | 527 | 27.51 | Uvigerina | -0.84 | 4.33 | 973-5-124 | 723 | 41.27 | Uvigerina | -1.52 | 3.72 |
| 973-4-52 | 547 | 29.83 | Uvigerina | -1.85 | 4.31 | 973-5-137 | 830 | 45.15 | Uvigerina | -1.31 | 1.79 |
| 973-4-54 | 567 | 32.14 | Uvigerina | -0.90 | 4.35 | 973-5-150 | 920 | 48.42 | Uvigerina | -1.72 | 4.30 |
| 973-4-56 | 587 | 34.45 | Uvigerina | -1.16 | 4.21 |
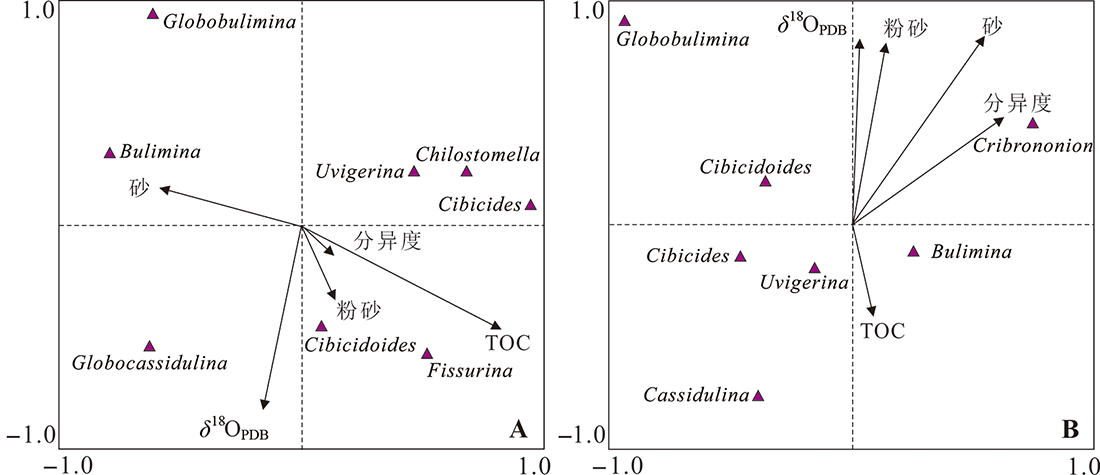
图10 973-4(A)和973-5(B)岩心主要底栖有孔虫类群丰度与环境因子(TOC、砂和粉砂含量,δ18OUvigerina spp.值)的CCA分析
Fig.10 CCA analysis of abundance of major benthic foraminifera taxa and environmental factors (contents of TOC, sand and silt, values of δ18OUvigerina spp.)for Cores 973-4 (A) and 973-5 (B)
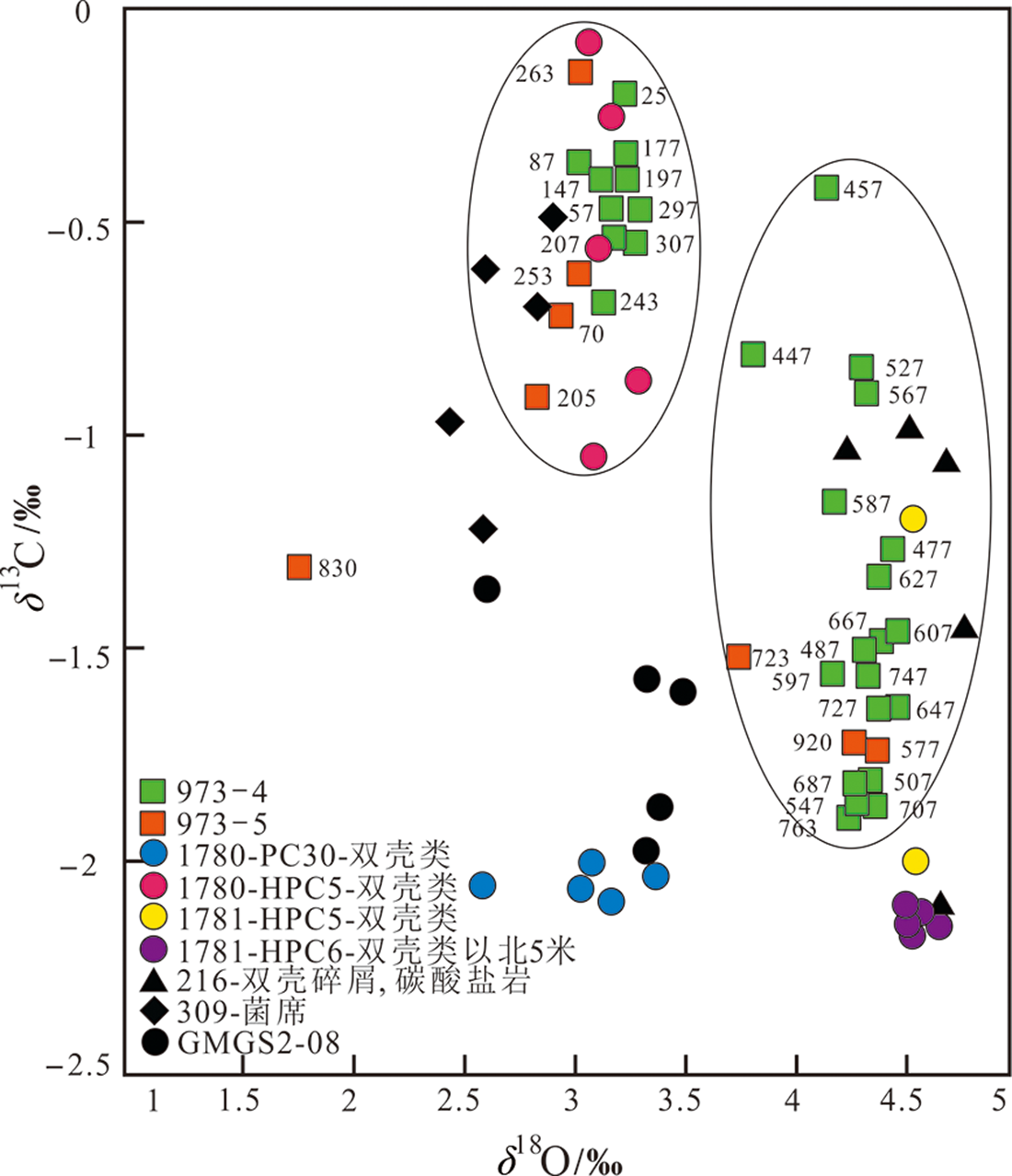
图11 973-4和973-5站Uvigerina壳体稳定氧碳同位素和典型冷泉微生境记录比较(Monterey湾1780和1781站位引自文献[60];Hikurangi Margin 216和309站位引自文献[61];南海东沙GMGS2-08站位引自文献[79])
Fig.11 Comparison of stable oxygen/carbon isotope and typical cold seep microhabitats of Uvigerina shells in Cores 973-4 and 973-5. Sites 1780 and 1781 in Monterey Bay adapted from [60]; Sites 216 and 309 in Hikurangi Margin adapted from [61]; Site GMGS2-08 in Dongsha area of South China Sea adapted from [79].
| 站位 | 微生境表征参数 | MIS 3 | MIS 2 | MIS 1 |
|---|---|---|---|---|
| δ18O聚类群 | >3.5‰类 | >3.5‰类/<3.5‰类 | <3.5‰类 | |
| 973-4 | 岩心深度/cm | 770.00~539.85 | 539.85~207.04 | 207.04~15.00 |
| 样品深度范围/cm与(相关微生境) | 770~547(1781站双壳,216站双壳碎屑和碳酸盐岩) | 307~243(309站菌席和1780站双壳) 527~447(1781站双壳,216站双壳碎屑和碳酸盐岩) | 207~25(309站菌席和1780站双壳) | |
| 优势生物类别 | U.peregrina | Bulimina,Cibicidoides | U.vadescens,Cibicides | |
| 类群组成和丰度差异 | 最大 | 较小 | 大 | |
| 冷泉微生境 | 与双壳和碳酸盐岩区相邻 | 晚期为双壳-菌席边缘区, 早期为双壳和碳酸盐岩区 | 双壳-菌席边缘区 | |
| 973-5 | 岩心深度/cm | 935.00~459.19 | 459.19~220.74 | 220.74~15.00 |
| 样品深度范围/cm与(相关微生境) | 935~557(1781站双壳,216站双壳碎屑和碳酸盐岩) | 263~253(309站菌席和1780站双壳) | 205(309站菌席,1780站双壳),70(309 站菌席) | |
| 优势生物类别 | Chilostomella+Globobulimina | Cibicidoides | Bulimina | |
| 类群组成和丰度差异 | 最大 | 较小 | 大 | |
| 冷泉微生境 | 与双壳和碳酸盐岩区紧邻 | 双壳-菌席边缘区 | 双壳-菌席边缘区 |
表5 研究区底栖有孔虫壳体同位素和微生境对比及综合分析(δ18O聚类结果见本文图11)
Table 5 Comparison of benthic foraminiferal shell isotopes in the study area and in typical cold seep microhabitats, with comprehensive analysis results (clustering result based on δ18O values is seen in Figure 11 of this paper)
| 站位 | 微生境表征参数 | MIS 3 | MIS 2 | MIS 1 |
|---|---|---|---|---|
| δ18O聚类群 | >3.5‰类 | >3.5‰类/<3.5‰类 | <3.5‰类 | |
| 973-4 | 岩心深度/cm | 770.00~539.85 | 539.85~207.04 | 207.04~15.00 |
| 样品深度范围/cm与(相关微生境) | 770~547(1781站双壳,216站双壳碎屑和碳酸盐岩) | 307~243(309站菌席和1780站双壳) 527~447(1781站双壳,216站双壳碎屑和碳酸盐岩) | 207~25(309站菌席和1780站双壳) | |
| 优势生物类别 | U.peregrina | Bulimina,Cibicidoides | U.vadescens,Cibicides | |
| 类群组成和丰度差异 | 最大 | 较小 | 大 | |
| 冷泉微生境 | 与双壳和碳酸盐岩区相邻 | 晚期为双壳-菌席边缘区, 早期为双壳和碳酸盐岩区 | 双壳-菌席边缘区 | |
| 973-5 | 岩心深度/cm | 935.00~459.19 | 459.19~220.74 | 220.74~15.00 |
| 样品深度范围/cm与(相关微生境) | 935~557(1781站双壳,216站双壳碎屑和碳酸盐岩) | 263~253(309站菌席和1780站双壳) | 205(309站菌席,1780站双壳),70(309 站菌席) | |
| 优势生物类别 | Chilostomella+Globobulimina | Cibicidoides | Bulimina | |
| 类群组成和丰度差异 | 最大 | 较小 | 大 | |
| 冷泉微生境 | 与双壳和碳酸盐岩区紧邻 | 双壳-菌席边缘区 | 双壳-菌席边缘区 |
| [1] |
ANDERSON R K, SCALAN R S, PARKER P L, et al. Seep oil and gas in gulf of Mexico slope sediment[J]. Science, 1983, 222(4624): 619-621.
DOI URL |
| [2] |
PAULL C K, HECKER B, COMMEAU R, et al. Biological communities at the Florida Escarpment resemble hydrothermal vent taxa[J]. Science, 1984, 226(4677): 965-967.
DOI URL |
| [3] | SUESS E, CARSON B, RITGER S, et al. Biological communities at vent sites along the subduction zone off Oregon[J]. Bulletin of the Biological Society of Washington, 1985, 6: 475-484. |
| [4] |
SIBUET M, OLU K. Biogeography, biodiversity and fluid dependence of deep-sea cold-seep communities at active and passive margins[J]. Deep Sea Research Part II, 1998, 45(1): 517-567.
DOI URL |
| [5] |
VALENTINE D L. Biogeochemistry and microbial ecology of methane oxidation in anoxic environments: a review[J]. Antonie Van Leeuwenhoek, 2002, 81(1/2/3/4): 271-282.
DOI URL |
| [6] | LEVIN L A. Ecology of cold seep sediments: interactions of fauna with flow, chemistry and microbes[C]//GIBSON R N, ATKINSON R J A, GORDON J D M. Oceanography and marine biology: an annual review. Boca Raton: CRC Press-Taylor & Francis Group, 2005, 43: 1-46. |
| [7] |
GRUPE B M, KRACH M L, PASULKA A L, et al. Methane seep ecosystem functions and services from a recently discovered southern California seep[J]. Marine Ecology, 2015, 36: 91-108.
DOI URL |
| [8] |
KVENVOLDEN K A. Gas hydrates-geological perspective and global change[J]. Reviews of Geophysics, 1993, 31(2): 173-187.
DOI URL |
| [9] | BEAUCHAMP B. Natural gas hydrates: myths, facts and issues[J]. Comptes Rendus Géoscience, 2004, 336(9): 751-765. |
| [10] |
SUESS E. Marine cold seeps and their manifestations: geological control, biogeochemical criteria and environmental conditions[J]. International Journal of Earth Sciences, 2014, 103(7): 1889-1916.
DOI URL |
| [11] |
FOUCHER J P, WESTBROOK G K, BOETIUS A, et al. Structure and drivers of cold seep ecosystems[J]. Oceanography, 2009, 22(1): 92-109.
DOI URL |
| [12] |
SEABROOK S, DE LEO F C, BAUMBERGER T, et al. Heterogeneity of methane seep biomes in the Northeast Pacific[J]. Deep-sea Research Part Ii-topical Studies in Oceanography, 2018, 150: 195-209.
DOI URL |
| [13] | SUESS E. RV Sonne cruise report SO177: SiGer 2004; Sino-German Cooperative Project; South China Sea continental margin: geological methane budget and environmental effects of methane emissions and gas hydrates[R]. Kiel, Germany, IFM-GEOMAR, 2005. |
| [14] | 黄永样, SUESS E, 吴能友, 等. 南海北部陆坡甲烷和天然气水合物地质: 中德全作SO-177航次成果专报[R]. 北京: 地质出版社, 2008. |
| [15] | 杨胜雄. 中国天然气水合物资源报告[R]// 中国地质调查局. 中国地质调查百项成果. 北京: 地质出版社, 2016: 432-440. |
| [16] |
FENG D, QIU J, HU Y, et al. Cold seep systems in the South China Sea: an overview[J]. Journal of Asian Earth Sciences, 2018, 168: 3-16.
DOI URL |
| [17] | GOODAY A J. Benthic foraminifera (Protista) as tools in deep-water palaeoceanography: environmental influences on faunal characteristics[J]. Advances in Marine Biology, 2003, 46: 1-90. |
| [18] | 向荣, 刘芳, 陈忠, 等. 冷泉区底栖有孔虫研究进展[J]. 地球科学进展, 2010, 25(2): 193-202. |
| [19] | 陈芳, 周洋, 刘广虎. 冷泉甲烷渗漏环境底栖有孔虫研究回顾与前景[J]. 海洋地质与第四纪地质, 2011, 31(2): 145-152. |
| [20] | MERINERO R, LUNAR R, CARDENES V, et al. Sunflower micro-pyrite in methane-derived carbonate pipes of the Gulf of Cadiz[J]. Resúmenes sobre el VIII Simposio MIA15, Málaga, Spain, 2015: 165-168. |
| [21] |
PANIERI G, LEPLAND A, WHITEHOUSE M J, et al. Diagenetic Mg-calcite overgrowths on foraminiferal tests in the vicinity of methane seeps[J]. Earth and Planetary Science Letters, 2017, 458: 203-212.
DOI URL |
| [22] |
SCHNEIDER A, CRÉMIÈRE A, PANIERI G, et al. Diagenetic alteration of benthic foraminifera from a methane seep site on Vestnesa Ridge (NW Svalbard)[J]. Deep Sea Research Part I: Oceanographic Research Papers, 2017, 123: 22-34.
DOI URL |
| [23] |
BORRELLI C, GABITOV R I, LIU M C, et al. The benthic foraminiferal δ 34S records flux and timing of paleo methane emissions[J]. Scientific Reports, 2020, 10(1): 1-10.
DOI URL |
| [24] | 陈芳, 苏新, 陆红锋, 等. 南海北部浅表层沉积底栖有孔虫碳同位素及其对富甲烷环境的指示[J]. 海洋地质与第四纪地质, 2007, 27(4): 1-7. |
| [25] | 陈芳, 周洋, 苏新, 等. 南海神狐海域含水合物层底栖有孔虫群落结构与同位素组成[J]. 海洋地质与第四纪地质, 2010, 30(2): 1-8. |
| [26] | 曹超, 雷怀彦. 南海北部有孔虫碳氧同位素特征与晚第四纪水合物分解的响应关系[J]. 吉林大学学报(地球科学版), 2012, 42(增刊1): 162-171. |
| [27] | 向荣, 方力, 陈忠, 等. 东沙西南海域表层底栖有孔虫碳同位素对冷泉活动的指示[J]. 海洋地质与第四纪地质, 2012, 32(4): 17-24. |
| [28] | 庄畅, 陈芳, 程思海, 等. 南海北部天然气水合物远景区末次冰期以来底栖有孔虫稳定同位素特征及其影响因素[J]. 第四纪研究, 2015, 35(2): 422-432. |
| [29] | 黄怡, 王淑红, 颜文, 等. 南海北部东沙海域天然气水合物分解事件及其与海底滑塌的关系[J]. 热带海洋学报, 2018, 37(4): 61-69. |
| [30] | 周洋, 陈芳, 苏新, 等. 南海东沙海域HD319岩心富甲烷环境底栖有孔虫群落结构[J]. 海洋地质与第四纪地质, 2009, 29(3): 5-12. |
| [31] | 杨艺萍, 唐灵刚, 向荣, 等. 东沙西南海域表层沉积物底栖有孔虫群落特征及其对冷泉活动的指示意义[J]. 微体古生物学报, 2017, 34(3): 237-246. |
| [32] | 杜德莉. 台西南盆地的构造演化与油气藏组合分析[J]. 海洋地质与第四纪地质, 1994, 14(3): 5-18. |
| [33] | 钟建强, 黄慈流. 台西南盆地晚新生代地质演化分析[J]. 海洋科学, 1994, (4): 34-38. |
| [34] | 徐尚, 王英民, 彭学超, 等. 台湾峡谷中段沉积特征及流体机制探讨[J]. 地质论评, 2013, 59(5): 845-852. |
| [35] | 罗祎, 苏新, 陈芳, 等. 南海北部DSH-1C柱状样晚更新世以来沉积物磁性特征及其环境意义[J]. 现代地质, 2010, 24(3): 521-527. |
| [36] | 张光学, 梁金强, 陆敬安, 等. 南海东北部陆坡天然气水合物藏特征[J]. 天然气工业, 2014, 34(11): 47-48. |
| [37] | 滕田田, 苏新, 刘浩东, 等. 南海东沙深海冷泉区973-5重力柱沉积物古菌多样性[J]. 现代地质, 2020, 34(1): 1-13. |
| [38] | LISIECKI L E, RAYMO M E. A Pliocene-Pleistocene stack of 57 globally distributed benthic δ18O records[J]. Paleoceanography, 2005, 20(1): PA1003. |
| [39] |
COHEN K M, GIBBARD P L. Global chronostratigraphical correlation table for the last 2.7 million years, version 2019 QI-500[J]. Quaternary International, 2019, 500: 20-31.
DOI URL |
| [40] | 李梦君, 毕乃双, 胡丽沙, 等. 南海北部台湾峡谷“蛟龙号”第140潜次沉积物特征及其沉积过程指示意义[J]. 海洋地质与第四纪地质, 2019, 39(4): 23-33. |
| [41] |
YIM W S, HUANG G, FONTUGNE M R, et al. Postglacial sea-level changes in the northern South China Sea continental shelf: evidence for a post-8200 calendar yr BP meltwater pulse[J]. Quaternary International, 2006, 145/146: 55-67.
DOI URL |
| [42] | 涂霞. 南海东北部海区有孔虫的分布及其与海洋环境的关系[J]. 热带海洋, 1983(1): 11-19. |
| [43] |
YIN J, LIU C, ZHANG J, et al. Distribution and constraining factors of planktonic and benthic foraminifers in bottom sediments of the southern South China Sea[J]. Palaeogeography, Palaeoclimatology, Palaeoecology, 2018, 502: 130-146.
DOI URL |
| [44] |
CORLISS B H, CHEN C. Morphotype patterns of Norwegian Sea deep-sea benthic foraminifera and ecological implications[J]. Geology, 1988, 16(8): 716-719.
DOI URL |
| [45] | KOHO K A, PIÑA-OCHOA E. Benthic foraminifera: inhabitants of low-oxygen environments[M]//ALTENBACH A, BERNHARD J, SECKBACH J. Anoxia. Cellular origin, life in extreme habitats and astrobiology. Dordrecht: Springer, 2012, 21: 249-285. |
| [46] | BÜHRING C, SARNTHEIN M, ERLENKEUSER H. Toward a high-resolution stable isotope stratigraphy of the last 1.1 my: Site 1144, South China Sea[J]. Proceedings of the Ocean Drilling Program: Scientific Results, 2006, 184(6): 1-29. |
| [47] | 张必东, 邬黛黛, 吴能友. 南海北部东沙海域沉积物地球化学特征及其反映的冷泉活动[J]. 海洋地质前沿, 2015, 31(9): 14-27. |
| [48] | MURRAY J W. Ecology and applications of benthic foraminifera[M]. Cambridge: Cambridge University Press, 2006: 238-264. |
| [49] | 卞云华, 王律江, 汪品先. 底栖有孔虫指示含氧量与古生产力: 南海北部陆坡晚第四纪的实例[M]//业治铮, 汪品先. 南海晚第四纪古海洋学研究. 青岛: 青岛海洋大学出版社, 1992: 227-233. |
| [50] |
KAIHO K. Effect of organic carbon flux and dissolved oxygen on the benthic foraminiferal oxygen index (BFOI)[J]. Marine Micropaleontology, 1999, 37(1): 67-76.
DOI URL |
| [51] |
PEARSON P N. Oxygen isotopes in foraminifera: overview and historical review[J]. Paleontological Society Papers, 2012, 18: 1-38.
DOI URL |
| [52] |
KUHNT W, HESS S, JIAN Z. Quantitative composition of benthic foraminiferal assemblages as a proxy indicator for organic carbon flux rates in the South China Sea[J]. Marine Geology, 1999, 156(1/2/3/4): 123-157.
DOI URL |
| [53] |
RISGAARD P N, LANGEZAAL A M, INGVARDSEN S, et al. Evidence for complete denitrification in a benthic foraminifer[J]. Nature, 2006, 443(7107): 93-96.
DOI URL |
| [54] |
GLOCK N, ROY A S, ROMERO D, et al. Metabolic preference of nitrate over oxygen as an electron acceptor in foraminifera from the Peruvian oxygen minimum zone[J]. Proceedings of the National Academy of Sciences, 2019, 116(8): 2860-2865.
DOI URL |
| [55] |
MACKENSEN A, SCHMIEDL G, Thiele J, et al. Microhabitat preferences of live benthic foraminifera and stable carbon isotopes off SW Svalbard in the presence of widespread methane seepage[J]. Marine Micropaleontology, 2017, 132: 1-17.
DOI URL |
| [56] | BOUCHET V M, TELFORD R J, RYGG B, et al. Can benthic foraminifera serve as proxies for changes in benthic macrofaunal community structure? Implications for the definition of reference conditions[J]. Marine Environmental Research, 2018: 24-36. |
| [57] |
VENTURELLI R A, RATHBURN A E, BURKETT A M, et al. Epifaunal foraminifera in an infaunal world: insights into the influence of heterogeneity on the benthic ecology of oxygen-poor, deep-sea habitats[J]. Frontiers in Marine Science, 2018, 5: 1-13.
DOI URL |
| [58] |
RATHBURN A E, CORLISS B H, TAPPA K D, et al. Comparisons of the ecology and stable isotopic compositions of living (stained) benthic foraminifera from the Sulu and South China Seas[J]. Deep Sea Research Part I: Oceanographic Research Papers, 1996, 43(10): 1617-1646.
DOI URL |
| [59] |
RATHBURN A E, LEVIN L A, HELD Z, et al. Benthic foraminifera associated with cold methane seeps on the northern California margin: ecology and stable isotopic composition[J]. Marine Micropaleontology, 2000, 38(3): 247-266.
DOI URL |
| [60] | MARTIN J B, DAY S A, RATHBURN A E, et al. Relationships between the stable isotopic signatures of living and fossil foraminifera in Monterey Bay, California[J]. Geochemistry, Geophysics, Geosystems, 2004, 5(4): Q04004. |
| [61] |
MARTIN R A, NESBITT E A, CAMPBELL K A. The effects of anaerobic methane oxidation on benthic foraminiferal assemblages and stable isotopes on the Hikurangi Margin of eastern New Zealand[J]. Marine Geology, 2010, 272(1/2/3/4): 270-284.
DOI URL |
| [62] |
BERNARD S, DAVAL D, ACKERER P, et al. Burial-induced oxygen-isotope re-equilibration of fossil foraminifera explains ocean paleotemperature paradoxes[J]. Nature Communications, 2017, 8(1): 1134.
DOI URL |
| [63] | HOOGAKKER B A, ELDERFIELD H, OLIVER K I, et al. Benthic foraminiferal oxygen isotope offsets over the last glacial-interglacial cycle[J]. Paleoceanography, 2010, 25(4): PA4229. |
| [64] | JIAN Z. Stable isotopic records of the glacial deep-water properties in the South China Sea[J]. Science in China Series D: Earth Sciences, 1998, 41(4): 337-344. |
| [65] |
DEJARDIN R, KENDER S, ALLEN C S, et al. “Live” (stained) benthic foraminiferal living depths,stable isotopes, and taxonomy offshore South Georgia, Southern Ocean: implications for calcification depths[J]. Journal of Micropalaeontology, 2018, 37(1): 25-71.
DOI URL |
| [66] |
BURKETT A, RATHBURN A E, PEREZ M E, et al. Influences of thermal and fluid characteristics of methane and hydrothermal seeps on the stable oxygen isotopes of living benthic foraminifera[J]. Marine and Petroleum Geology, 2018, 93: 344-355.
DOI URL |
| [67] |
NAEHR T H, EICHHUBL P, ORPHAN V J, et al. Authigenic carbonate formation at hydrocarbon seeps in continental margin sediments: a comparative study[J]. Deep-sea Research Part Ii-topical Studies in Oceanography, 2007, 54(11): 1268-1291.
DOI URL |
| [68] |
BOHRMANN G, GREINERT J, SUESS E, et al. Authigenic carbonates from the Cascadia subduction zone and their relation to gas hydrate stability[J]. Geology, 1998, 26(7): 647-650.
DOI URL |
| [69] |
GUPTA B K, AHARON P. Benthic foraminifera of bathyal hydrocarbon vents of the Gulf of Mexico: initial report on communities and stable isotopes[J]. Geo-marine Letters, 1994, 14(2): 88-96.
DOI URL |
| [70] | WEFER G, HEINZE P M, BERGER W H, et al. Clues to ancient methane release[J]. Nature, 1994, 369(6478): 282. |
| [71] |
AHARON P, FU B. Microbial sulfate reduction rates and sulfur and oxygen isotope fractionations at oil and gas seeps in deepwater Gulf of Mexico[J]. Geochimica et Cosmochimica Acta, 2000, 64(2): 233-246.
DOI URL |
| [72] |
WORTMANN U G, CHERNYAVSKY B, BERNASCONI S M, et al. Oxygen isotope biogeochemistry of pore water sulfate in the deep biosphere: dominance of isotope exchange reactions with ambient water during microbial sulfate reduction (ODP Site 1130)[J]. Geochimica et Cosmochimica Acta, 2007, 71(17): 4221-4232.
DOI URL |
| [73] |
HESSE R, HARRISON W E. Gas hydrates (clathrates) causing pore-water freshening and oxygen isotope fractionation in deep-water sedimentary sections of terrigenous continental margins[J]. Earth and Planetary Science Letters, 1981, 55(3): 453-462.
DOI URL |
| [74] |
DAVIDSON D W, LEAIST D G, HESSE R. Oxygen-18 enrichment in the water of a clathrate hydrate[J]. Geochimica et Cosmochimica Acta, 1983, 47(12): 2293-2295.
DOI URL |
| [75] |
USSLER W, PAULL C K. Effects of ion exclusion and isotopic fractionation on pore water geochemistry during gas hydrate formation and decomposition[J]. Geo-marine Letters, 1995, 15(1): 37-44.
DOI URL |
| [76] | MATSUMOTO R, BOROWSKI W S. Gas hydrate estimates from newly determined oxygen isotopic fractionation (αGH-IW) and δ18O anomalies of the interstitial waters: Leg 164, Blake Ridge[J]. Proceedings of the Ocean Drilling Program: Scientific Results, 2000, 164: 59-66. |
| [77] | KANO A, MIYAHARA R, YANAGAWA K, et al. Gas hydrate estimates in muddy sediments from the oxygen isotope of water fraction[J]. Chemical Geology, 2017(8): 107-115. |
| [78] | MACKENSEN A, WOLLENBURG J E, LICARI L, et al. Low δ 13C in tests of live epibenthic and endobenthic foraminifera at a site of active methane seepage[J]. Paleoceanography, 2006, 21(2): PA2022. |
| [79] |
WAN S, FENG D, CHEN F, et al. Foraminifera from gas hydrate-bearing sediments of the northeastern South China Sea: proxy evaluation and application for methane release activity[J]. Journal of Asian Earth Sciences, 2018, 168: 125-136.
DOI URL |
| [80] |
TRÉHU A M, BOHRMANN G, RACK F R, et al. Proceedings of the Ocean Drilling Program, Initial Reports.Volume 204, Leg 204 summary-Ocean Drilling Program[R]. DOI: 10.2973/odp.proc.ir.204.2003.
DOI |
| [81] |
HILL T M, KENNETT J P, VALENTINE D L. Isotopic evidence for the incorporation of methane-derived carbon into foraminifera from modern methane seeps, Hydrate Ridge, Northeast Pacific[J]. Geochimica et Cosmochimica Acta, 2004, 68(22): 4619-4627.
DOI URL |
| [82] |
SAHLING H, RICKERT D, LEE R W, et al. Macrofaunal community structure and sulfide flux at gas hydrate deposits from the Cascadia convergent margin, NE Pacific[J]. Marine Ecology Progress Series, 2002, 231: 121-138.
DOI URL |
| [83] |
HEINZ P, SOMMER S, PFANNKUCHE O, et al. Living benthic foraminifera in sediments influenced by gas hydrates at the Cascadia convergent margin, NE Pacific[J]. Marine Ecology Progress Series, 2005, 304: 77-89.
DOI URL |
| [84] | 于晓果, 韩喜球, 李宏亮, 等. 南海东沙东北部甲烷缺氧氧化作用的生物标志化合物及其碳同位素组成[J]. 海洋学报, 2008, 30(3): 77-84. |
| [1] | 窦衍光, 李清, 吴永华, 赵京涛, 孙呈慧, 蔡峰, 陈晓辉, 张勇, 范佳慧, 石学法. 冲绳海槽MIS6期以来底栖有孔虫碳氧同位素特征及其古海洋指示意义[J]. 地学前缘, 2022, 29(4): 84-92. |
| [2] | 袁洁琼, 丁旋, 邹欣庆. 南黄海辐射沙脊群表层沉积物中底栖有孔虫埋葬群分布特征及其环境意义[J]. 地学前缘, 2020, 27(6): 276-288. |
| [3] | 梁劲,王静丽,陆敬安,康冬菊,匡增桂,杨承志. 台西南盆地含天然气水合物沉积层测井响应规律特征及其地质意义[J]. 地学前缘, 2017, 24(4): 32-40. |
| [4] | 梁静之, 黄宝琦, 董轶婷, 贾文博, 周彦希. 南海北部MD12-3432站MIS 11期以来底栖有孔虫反映的古环境变化[J]. 地学前缘, 2016, 23(4): 292-300. |
| 阅读次数 | ||||||
|
全文 |
|
|||||
|
摘要 |
|
|||||